Capire la Resistenza
Prof. Salvatore Buzzelli
Introduzione
La RESISTENZA, generalmente considerata, identifica la qualità fisica che permette ad un soggetto o ad un distretto biomeccanico, di protrarre uno sforzo nel tempo.
Esclusi i movimenti che richiedono un numero limitato di contrazioni muscolari, che quindi si esauriscono nel volgere di pochi secondi, tutti gli sforzi che si protraggono oltre i 15”- 20” richiedono doti di resistenza..
Nella pratica, si è soliti indicare l’ intensità e la durata dello sforzo per poter fare una distinzione più specifica della Resistenza , infatti l' intensità dello sforzo richiesto e la sua durata, sono i parametri che influenzeranno la denominazione, sistemi ed apparati coinvolti e la metodologia di allenamento più idonea per creare i presupposti dell' adattamento desiderato.
Si considererà allora se ci si riferisce a sforzi di media durata, che durano fino a circa 2' ad intensità molto elevata, o a sforzi di lunga durata che vanno dai 3', a qualche ora ad intensità decrescente da elevata a blanda.
I prerequisiti legati alla resistenza, per sperare in risultati agonistici di livello, spaziano in più settori: da quelli relativi alla struttura intima del muscolo e più precisamente alla percentuale di fibre di tipo ossidativo in esso presente (che deve essere alta), alla capacità dei tessuti di utilizzare l’ ossigeno trasportato dal sangue per mezzo dell’ emoglobina, dalla capacità del sistema cardiovascolare di far arrivare rapidamente gli elementi nutritivi ai distretti biomeccanici impegnati nell’ attività, alla funzionalità di sistemi deputati alla eliminizione delle scorie rilasciate dal lavoro muscolare, e ancora una quantità considerevole di fattori di tipo organico e psicologici.
Prima di addentrarci nello studio della Resistenza e nel campo della metodologia dell'allenamento ad essa relativa, bisogna avere ben chiare le nozioni inerenti la struttura anatomofisiologica del muscolo e le reazioni biochimiche fondamentali, responsabili della produzione di energia per il lavoro muscolare.
Per questo motivo, prima di esporre le metodologie allenanti e i fini che queste si prepongono, vale la pena di fare una rivisitazione in chiave specifica relativa allo sport dei sistemi anatomofisiologici e bioenergetici che intervengono nella resistenza.
FONTI ENERGETICHE E LAVORO MUSCOLARE
Cercheremo di sintetizzare il funzionamento delle FONTI ENERGETICHE, descrivendone i principi di attivazione e le caratteristiche del loro ripristino.
Sappiamo che per dar luogo ad un movimento, bisogna attivare l’ apparato locomotore, e che i muscoli che presiedono ad un determinato movimento debbano disporre di energia. Questa è immagazinata sotto forma di legami chimici ad alto potenziale in un nucleotide chiamato ATP (acido Adenosintrifosforico). L’ ATP, che si trova depositato nei muscoli in minime quantità, dopo aver liberato l’energia per l’ attivazione muscolare, deve essere continuamente ricostituito in quanto solo l’ ATP è utilizzabile dai muscoli come combustibile.
Il compito di ricaricare l’ ATP, allora viene assolto da fonti energetiche collaterali, il cui compito consiste nel mettere a disposizione i rispettivi substrati energetici e sistemi enzimatici, al fine di tenere sempre carico il serbatoio di energia utile per la contrazione muscolare, che ricordiamo e' sempre e solo l'ATP.
Possiamo schematizzare nella seguente tabella le reazioni responsabili della produzione di energia.
E' consequenziale capire che più substrati organici sono disponibili, più a lungo è possibile riformare ATP; non solo, ma anche che se la richiesta energetica è relativa a sforzi molto intensi, diviene oltremodo importante che i tempi nei quali i substrati vengono trasformati in energia di resintesi, siano necessariamente brevi.
Quindi per ogni processo energetico si identificano la CAPACITA’ e la POTENZA.
La CAPACITA' rappresenta la quantità di substrato disponibile, mentre la quantità di substrato trasformato in energia di resintesi nell' unita' di tempo, viene chiamata POTENZA della fonte energetica.
Le fonti energetiche a loro volta si dividono in:
- AEROBICA ( reazione C dello schema; per esercitazioni medie di lunga durata)
- ANAEROBICA ALATTACIDA (reazione A ; per esercitazioni esplosive)
- ANAEROBICA LATTACIDA (reazione D ; per esercitazioni intense di breve durata)
Nel caso della resistenza interesserà dunque comprendere i meccanismi bioenergetici della fonte Aerobica e della fonte Anaerobica Lattacida; quindi approfondiremo i concetti di Capacità e Potenza Aerobica, e di Capacità e Potenza Lattacida a seconda che le fonti energetiche stimolate, siano esse Aerobica o Lattacida.
EFFETTO DELL’ ACIDO LATTICO SUL LAVORO MUSCOLARE
L’Acido Lattico (LA) è un metabolita del muscolo derivante dall’ idrolisi anaerobica del glicogeno.Si forma per fermentazione dell’ Acido Piruvico in presenza dell’ enzima specifico LATTICODEIDROGENASI.
E’ stato dimostrato che basse concentrazioni di LA si formano già a partire dall’ inizio di una qualsiasi attività fisica, anche se la sua presenza massiccia è determinata dall’ esecuzione di un esercizio strenuo che duri almeno 30- 40 secondi.
La presenza di questa sostanza, altera l’ ambiente cellulare dove il sarcomero è impegnato ad operare, rendendo difficile la contrazione muscolare, di conseguenza riducendo l’ efficienza del muscolo fino a renderne impossibile la continuazione del lavoro.
Infatti gli ioni idrogeno (H+) dell’ acido lattico, antagonizzando gli ioni calcio (Ca++), la cui presenza nel sarcomero favoriscono la realizzazione dei ponti actomiosinici, inibiscono progressivamente nel corso di una esercitazione strenua o comunque di medio-alta intensità, la contrazione muscolare fino all’ arresto vero e proprio dell’ attività.
L’ acido lattico prodotto, alla cessazione del lavoro muscolare, durante la fase di recupero, diviene un precursore del glicogeno, infatti viene riconvertito in tale sostanza percorrendo a ritroso la reazione che ne
ha determinato la sua formazione (Ciclo di Cori).
Le reazioni in questione avvengono in assenza di Ossigeno e per tale motivo sono dette Anaerobiche.
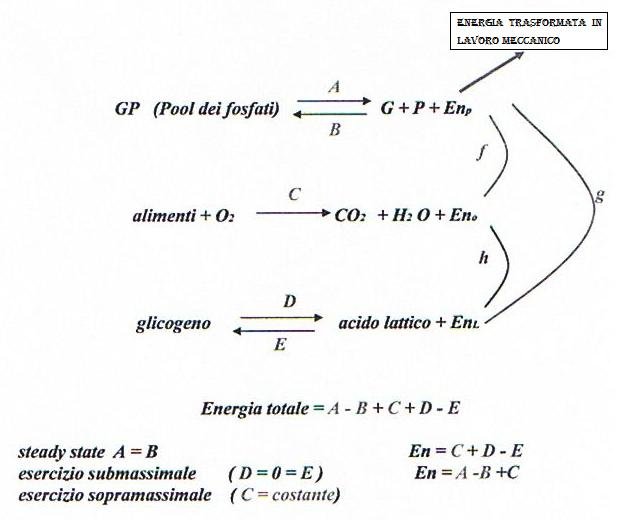
MODELLO IDRAULICO DEI MECCANISMI BIO- ENERGETICI DEL MUSCOLO
La fig. 1 rappresenta un modello idraulico delle fonti energetiche che hanno luogo nel muscolo.
Il liquido del serbatoio chiamato Fosfageno (pool dei fosfati :ATP, CP) comunica direttamente con l’ esterno attraverso il rubinetto T che regola il flusso della spesa energetica, il serbatoio Ossigeno rappresenta la fonte aerobica, e il serbatoio Acido Lattico rappresenta la fonte anaerobica lattacida. I serbatoi comunicano tra loro attraverso dei tubi in maniera da costituire un sistema di vasi comunicanti.
Quando il rubinetto T chiude il flusso si ha inattività muscolare e quindi non si ha spesa energetica. Il liquido nel sistema a vasi comunicanti è allo stesso livello.
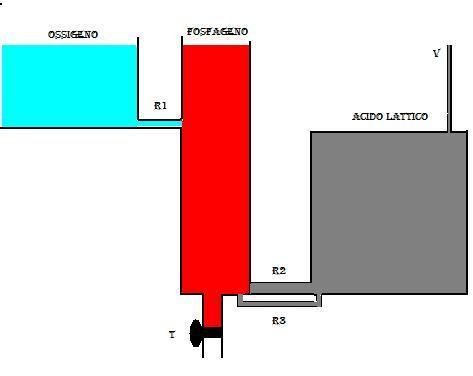
fig.1
Se si apre il rubinetto T (spesa energetica) (fig.2), il liquido GP defluisce all’ esterno e produce un abbassamento del livello nel serbatoio; dal serbatoio O2 il liquido passerà in quello GP attraverso il collegamento R1.
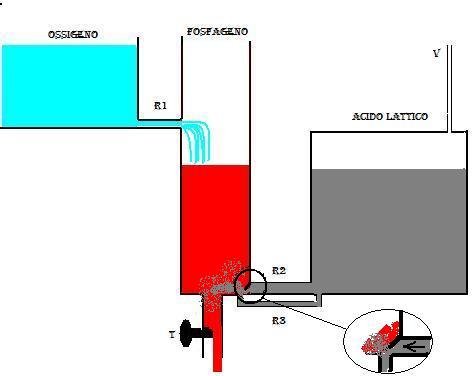
fig.2
La resistenza che oppone il tubo R1 è maggiore di quella offerta dal rubinetto t; se la spesa energetica è contenuta, allora dopo un certo periodo di tempo si avrà un assestamento del flusso tale che quello del liquido che defluisce dal serbatoio O2 corrisponde a quello che esce dal rubinetto t. Questo flusso in pratica rappresenta il consumo di ossigeno (VO2) nella condizione chiamata STEADY STATE.
Quando invece il rubinetto t è molto aperto, cioè si ha una notevole spesa energetica (nel caso di un esercizio sovramassimale), il serbatoio GP tende a svuotarsi rapidamente, e il liquido del serbatoio O2 non è sufficiente a rimpiazzare quello che esce dal serbatoio GP, in quanto anche se il serbatoio O2 ha capacità illimitata, la resistenza offerta dal tubo R1 è grande. Quando il livello del serbatoio GP scende sotto il livello dello sbocco della resistenza R1 allora la pressione presente nel serbatoio LA esercita una spinta sulla valvola del tubo R2 e fa sì che questa si apra verso il sebatoio GP facendo così defluire in esso il liquido contenuto in LA (fig.3)
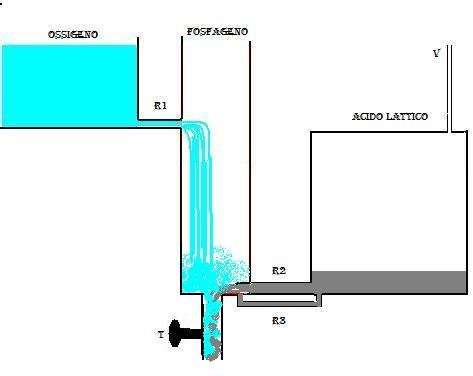
fig.3
Questo liquido rappresenta la componente di energia lattacida . In questo modo il livello del liquido in GP può essere mantenuto costante al di sotto dello sbocco R1, ma ciò non può durare molto tempo data la limitata capacità del serbatoio LA. Il flusso di liquido in R2 rappresenta la potenza del meccanismo lattacido.
Quando la massima apertura del rubinetto t ha condotto allo svuotamento del serbatoio GP, si chiude il rubinetto t e la spesa energetica si arresta.
Il serbatoio GP comincia a riempirsi per effetto del flusso che arriva da R1, e quando il livello GP raggiunge lo sbocco R1 il flusso diminuirà in maniera esponenziale fino al riempimento totale del serbatoio GP.
Se il rubinetto t viene chiuso prima che il serbatoio LA si sia svuotato completamente, nella prima fase del riempimento di GP ci sarà un contributo in liquido anche dal serbatoio LA fino a che la pressione presente in GP non equiparerà quella presente in LA, determinando la chiusura della valvola in R2.
Il riempimento di GP in questo caso avverrà più velocemente perchè sono due i serbatoi che concorrono.
Il serbatoio LA, intanto svuotato si riempirà usufruendo del liquido che riuscirà a passare attraverso la conduttura R3, che essendo molto stretta, oppone una grande resistenza e ritarderà alquanto il riempimento dl serbatoio LA.
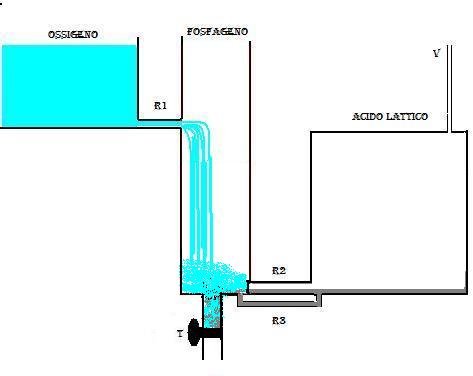
fig.4
Ciò che abbiamo descritto costituisce un perfetto modello analogico di quello che avviene nella realtà.
La fig. 5 mostra la ripartizione dei substrati produttori di energia nell’ erogazione energetica (Keul).
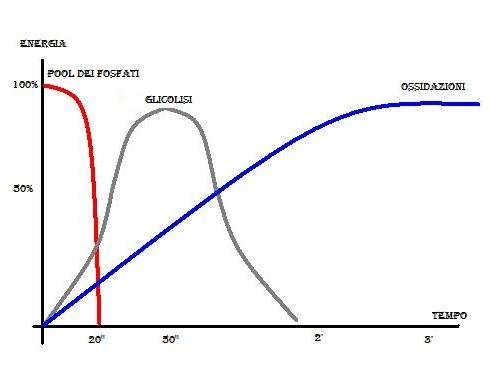
fig. 5
In un intenso sforzo fisico vengono intaccati dapprima i depositi di ATP. Essi possono procurare energia soltanto per un brevissimo tempo, ma con l’ esaurimento del deposito di ATP vengono intaccati anche i depositi di Creatinfosfato.Con i deopsiti dei fosfati ricchi di energia (pool dei fosfati), si possono raggiungere al massimo 20” di lavoro intenso. Con l’ inizio dello sforzo vengono tuttavia sollecitate anche le richieste energetiche a carico della glicolisi. Essa raggiunge il suo massimo dopo 40”-50” e poi viene sempre meno interessata all’ erogazione energetica. Le erogazioni ossidative prendono sempre più parte e diventano, infine, la fonte fondamentale ed esclusiva dell’ energia nel lavoro muscolare.
LE FIBRE MUSCOLARI
Nella struttura del muscolo esistono Fibre Lente e Fibre Veloci, la differenziazione tra esse è data oltre che dalla differenza di colorazione dovuta a presenza di sostanze caratterizzanti, e dal tipo di innervazione che ne determina la morfologia, soprattutto dalla differenza di concentrazione di enzimi capaci di attivare la liberazione di energia dall' ATP (BARANY, 1967). Le fibre lente hanno una minore concentrazione di ATPasi (enzima specifico dell' ATP) rispetto alle fibre veloci, ma di contro sono ricche di mioglobina, che essendo di colore rosso, ne determina anche l' appellativo di "Fibre Rosse".
Più precisamente, le fibre presenti nel muscolo scheletrico umano, sono di tre
tipi:
- le fibre tipo IA, dette anche: fibre lente, rosse, ossidative, o anche Slow Twich; queste riescono a sviluppare tensioni muscolari basse, ma sono capaci di protrarre la loro azione per un tempo molto lungo. Sono innervate da nervi di piccola dimensione e sono ricche di mitocondri.
- le fibre tipo IIA, dette anche fibre intermedie, (o anche fibre glicolitiche-ossidative); queste hanno la caratteristica di potersi comportare o come fibre lente o come quelle veloci, e principalmente subiscono gli adattamenti imposti dall' allenamento. Contengono sia gli enzimi della via glicolitica sia quelli tipici della via aerobica. Insomma sono una via di mezzo tra le fibre veloci e le fibre lente che possiamo considerare come fibre veloci-resistenti.
- le fibre tipo IIB, dette anche: fibre veloci, bianche, glicolitiche, o anche Fast Twich; queste sono innervate da nervi di grossa dimensione e forniscono tensioni di alta intensità ma essendo facilmente affaticabili, esauriscono il loro intervento in breve tempo. Esse sono ricche di ATPasi. L' insieme del nervo motore e le fibre che da esso vengono attivate, viene chiamato UNITÀ MOTRICE.
Non è ben chiaro se è il tipo di fibra che determina le dimensioni del nervo che le attiva, o il contrario; sta di fatto che se il nervo è grosso, innerva fibre veloci (della dimensione di 12-20 micron) e l' insieme motoneurone-fibre viene detto Unità Motoria Fasica.
Se il nervo è piccolo, innerva fibre lente (della dimensione di 6-12 micron) e l' insieme motoneurone-fibre viene detta Unità Motoria Tonica.
Le unità toniche solitamente sono formate dalle fibre dei muscoli che lavorano contro gravità e lavorano a bassa frequenza di stimoli tra i 20 e i 30 al secondo (20-30 Hertz); le unità fasiche lavorano a frequenze di stimolo superiori, dai 40 agli 80 Hz.
L' APPARATO CARDIOVASCOLARE
In linea generale,la funzione dell'apparato cardiovascolare e' quella garantire all'organismo le sostanze necessarie per il normale svolgersi dei processi metabolici cellulari, e di assumere i metaboliti formatisi dai suddetti processi per riversarli negli apparati preposti per la loro eliminazione.
Anatomicamente esso e' composto dal CUORE e dal sistema ARTERO-VENOSO che a loro volta per la loro differente struttura sono deputati ad assolvere differenti funzioni; precisamente il CUORE che deve fornire l' energia necessaria perche' sia assicurata la circolazione del sangue, le ARTERIE DI GROSSO CALIBRO che hanno la tunica media di struttura prevalentemente plastica svolgono il duplice compito di far defluire il sangue dai ventricoli e di trasformare il flusso sanguineo da intermittente a continuo; le ARTERIE DI MEDIO E PICCOLO CALIBRO che hanno il compito di regolare il flusso distrettuale nei vari organi (le metarteriole favorite dalla tunica media a struttura muscolare - sfinteri precapillari); i CAPILLARI la cui struttura e' molto semplice essendo forniti di una parete molto sottile praticamente formata da un unico tratto endoteliale che poggia su una membrana basale, il cui compito è quello di permettere uno scambio per diffusione tra il sangue ed il liquido interstiziale dei vari tessuti (e viceversa); le VENE che assicurano il ritorno del sangue all' atrio destro per effetto di un gradiente pressorio che è differente nei diversi circoli: nel circolo arterioso diminuisce in senso centrifugo, nel circolo venoso diminuisce in senso centripeto.
Istologicamente il CUORE e' costituito da due tipi di tessuto:
- il tessuto miocardico comune (tessuto muscolare vero e proprio) cui spetta la funzione meccanica della contrazione.
- il tessuto specifico che forma il sistema di conduzione atrio-ventricolare (tessuto istologicamente diverso da quello del miocardio comune) il cui compito e' quello di generare l' impulso e di condurre l' eccitamento dalla muscolatura atriale a quella ventricolare determinando la contrazione degli atrii prima e dei ventricoli poi.
Le formazioni che costituiscono il sistema di conduzione atrio-ventricolare sono:
- nodo del seno (Keith e Flack)
- nodo atrio-ventricolare (Tawara e Ashoff)
- fascio di His
- branca destra e sinistra del fascio di His
- rete del Purkinje
Le formazioni decorrono in senso cranio-caudale nel miocardio a partire dall'atrio destro, passando nel setto interatriale, poi nel setto interventricolare per diramarsi nell' endocardio ventricolare. Ogni formazione e' capace di elaborare ed emettere stimoli per la contrazione del cuore (automatismo del cuore) ma la sede piu' importante e' il nodo del seno. E' per questo motivo che in condizioni fisiologiche il ritmo cardiaco viene detto RITMO SINUSALE.
Il CICLO CARDIACO e' formato dalla SISTOLE e dalla DIASTOLE e mediamente dura 800 millisecondi.
La Diastole e' relativa al riempimento delle cavita' e all' irroramento del tessuto miocardico per conto del circolo CORONARICO, ed è la fase piu' lunga del ciclo cardiaco (all' incirca 500-600 millisecondi).
La Sistole e' la fase di eiezione del sangue dal cuore alla periferia.
Durante la Sistole Atriale si ha la Diastole Ventricolare e viceversa.
Tra le funzioni emodinamiche del cuore distinguiamo:
- GITTATA SISTOLICA, il volume di sangue che esce da ciascun ventricolo ad ogni contrazione ( da 70 a 100 mml a riposo)
- PORTATA CARDIACA o GITTATA CARDIACA o VOLUME/MINUTO il volume di sangue che esce da ciascun ventricolo in un minuto (da 5000 a 7000 mml di sangue al minuto a riposo)
La legge di MAESTRINI-STARLING
L' energia della contrazione muscolare e' in funzione della lunghezza delle fibre muscolari.Cioe' la quantita' di energia che si libera in una contrazione del muscolo cardiaco aumenta quando per un aumento del volume diastolico del cuore, si determina una maggior lunghezza delle fibre miocardiche (prestiramento o precarico).
E' importante assimilare questa nozione in quanto la ritroveremo negli aspetti degli adattamenti emodinamici del cuore dovuti all' allenamento.
IL MASSIMO CONSUMO DI OSSIGENO (VO2max)
Il Massimo Consumo di Ossigeno o VO2max è comunemente considerato il fattore determinante della prestazione di resistenza. In pratica rappresenta la quantità massima di ossigeno utilizzata dai muscoli attivi per la produzione di energia per il lavoro. Nella fig. 6 vediamo la differenza del VO2max tra Uomini e Donne, con un netto prevalere dei primi sulle seconde sia a livello normale, sia dopo l'allenamento sia in atleti evoluti e specializzati come i maratoneti.
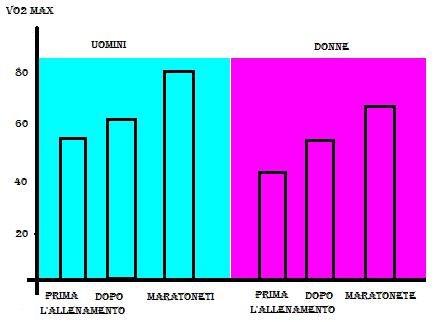
fig. 6
Il VO2max è determinato dal prodotto della gittata cardiaca per la differenza artero-venosa di ossigeno.
VO2max = (GS x FC) x diff. a-v O2
dove:
GS = gittata sistolica
FC = frequenza cardiaca
(GS x FC) = gittata cardiaca
La differenza atero-venosa di ossigeno è la conseguenza di due fattori:
1) la quantità di ossigeno che arriva al tessuto muscolare ( più ne viene prelevato, meno ne resterà nel sangue venoso, con conseguente aumento della differnza artero-venosa di ossigeno)
2) la distribuzione generale dl flusso sanguineo (capillarizzazione)
L’ unità di misura del VO2max è : litri di ossigeno al minuto o anche litri/minuto/kg.
Durante un’ attività di resistenza, non è solo l’ entità di VO2max ad essere importante, ma anche la sua percentuale di utilizzazione.
VO2max % = (VO2 / VO2max) x 100
Se per esempio un esercitazione richiede 1,5 litri/minuto di ossigeno (VO2) ed il VO2max è di 3,5 l/m, in tal caso si utilizzerebbe il 50% del VO2max.
Diversamente in base allo stesso consumo di ossigeno richiesto, ma con VO2max di 5 l/m, la percentuale di utilizzo risulterà del 30%. In altre parole, in quest’ ultimo caso l’ esercizio sarebbe più facile a parità di VO2.
Il VO2max % è utile come indice della pesantezza di un esercizio in rapporto alla massima capacità individuale.
Il VO2max è influenzato positivamente dalla presenza nei muscoli di una alta percentuale di fibre più adatte a sostenere un lavoro protratto nel tempo, cioè quelle di tipo IA, dette anche rosse o lente, e in conseguenza di ciò dalla massiccia presenza nei muscoli di elementi capaci di favorire la produzione prolungata di energia per il lavoro: i mitocondri.
I MITOCONDRI E LA RESPIRAZIONE CELLULARE
Sappiamo che le cellule e quindi gli organismi da esse formati necessitano continuamente di energia per mantenersi in vita.
Tale energia e' prodotta principalmente dalla demolizione del glucosio, dato che anche le reazioni metaboliche di altre sostanze dipendono strettamente da essa.
E' inoltre necessario che l' energia, che si forma da queste reazioni, possa venire immagazzinata per essere spesa quando richiesta; di ciò si incarica una molecola particolare, e precisamente un nucleotide, strutturata in modo da assorbire l' energia imprigionandola in un legame; questo legame potrà essere spezzato al momento opportuno rendendo così disponibile tale energia.
L'ultimo gruppo fosforico si stacca se occorre energia e si ottiene così l' ADP.
Questa molecola, quando viene prodotta energia, lega a sè un gruppo fosforico preso dalle riserve della cellula, immagazzinando in tale legame l' energia neoformata. La demolizione del glucosio avviene nel citoplasma: in parte nella frazione non strutturata di esso e in parte in organelli specializzati detti MITOCONDRI.
I mitocondri, corpiccioli di varia forma generalmente più o meno allungata, hanno la parete esterna di costituzione analoga alla membrana cellulare mentre la parete interna è ricca di creste ed avvallamenti che ne aumentano la superficie: in tal modo può contenere più enzimi. I mitocondri sono gli organi della respirazione cellulare che avviene bruciando glucosio in presenza di ossigeno.
C6 H12 O6 + O6 > 6CO2 + 6H2O + 690000 cal.
Da tale reazione si ottengono 690.000 calorie per ogni mole di glucosio (pari a grammi 180). La scissione del glucosio avviene attraverso una serie di reazioni che possiamo distinguere in due gruppi fondamentali: il primo avviene in assenza di ossigeno, e' detto perciò GLICOLISI ANAEROBICA e si svolge nella parte del citoplasma con l' ausilio di enzimi qui localizzati.
La seconda fase detta AEROBIA , perchè avviene con intervento di O2, si svolge nei mitocondri e consta di due processi: il ciclo di Krebs e la catena citocromica. Perchè il glucosio possa essere utilizzato, deve essere attivato con H3 PO4, ciò provoca un dispendio di energia che però è largamente compensata con l'energia prodotta. La fosforilazione avviene nel C 1 e 6 e tale composto ha 6 atomi di carbonio si scinde in due molecole di 3 atomi di carbonio ciascuna che, attraverso successive trasformazioni, diventano ACIDO PIRUVICO (CH3-CO-COOH).
A questo punto, se l' O2 è sufficiente, l' acido piruvico va incontro a trasformazioni che permettono la sua ulteriore demolizione. Esso infatti reagisce con il Coenzima A (HSCoA) tramite un gruppo sulfidrilico qui messo in evidenza e con un coenzima deidrogenante DPN (NAD), interviene nella reazione un enzima decarbossilante che permette la liberazione del CO2.
CH3-CO-COOH +HSCoA + DPN > CH3-Co > SCoA +DPNH2+CO2
L'anidride carbonica viene espulsa, invece l'acetato attivo e il DPNH2 verranno utilizzati. Queste reazioni si erano svolte fuori dai mitocondri mentre ora queste sostanze penetrano all' interno.
Dentro ai mitocondri c' è una specie di "fuoco continuo" sempre in funzione formato da una serie di reazioni catalizzate da enzimi di diverso tipo che formano un ciclo detto di Krebs dove il primo composto (acido citrico) ha 6 atomi di carbonio, mentre l' ultimo (acido ossalacetico) ne ha 4, in quanto durante il ciclo per due volte viene espulsa CO2. L' ultimo composto si lega all' acido acetico ottenuto dalla reazione:
CH3-CO SCoA + H2O > CH3-COOH + HSCoA
che con i suoi due atomi di carbonio lo ritrasforma nell' acido citrico originario.
Così le reazioni ricominciano ed il CH3-COOH viene consumato perchè viene espulsa CO2.
Durante il ciclo viene espulso inoltre H2, che però non viene scartato, ma caricato da altri DPN. Il DPNH2 scarica H2 ad altri trasportatori (FAD) fino ai citocromi che sono una serie di molecole proteiche coniugate il cui gruppo prosteico contiene Fe.
Il Fe puo' essere bivalente o trivalente perchè possiede il livello interno pseudosaturo
Quando l'H2 arriva alla catena dei citocromi i suoi protoni si separano dagli elettroni: i protoni restano fuori dalla catena citocromica che non li accetta, l' elettrone viene passato da un citocroma all'altro ( ovviamente per ogni H2 occorrono due citocromi con un atomo di Fe ciascuno) infatti i citocromi contengono Fe+++ per cui caricando un H- diventa Fe++ poi passandolo al citocroma successivo tornano come prima. Lungo questa catena gli H- spostandosi perdono energia che viene immagazzinata nell' ATP, una volta scarichi vengono ceduti all' O2 della respirazione che diventa O--.
Le due cariche negative attirano i due protoni H+ formando l' acqua che verrà usata dalla cellula secondo le necessità oppure espulsa.
MODIFICAZIONI DELLA CONTRATTILITA' MIOCARDICA NEL LAVORO MUSCOLARE
L' esecuzione di una attività muscolare di un certo impegno, per l' economia dell' organismo, induce necessariamente l' intervento di fenomeni emodinamici che riescono a supplire alle aumentate esigenze metaboliche dei distretti muscolari impegnati nel lavoro stesso. Non solo, ma tali modificazioni devono garantire di mantenere per un certo tempo la nuova condizione dell' organismo e di riequilibrare le eventuali alterazioni dei sistemi metabolici e fornitori di energia, una volta cessato il lavoro muscolare.
Quindi, all' inizio di una prestazione muscolare di un certo impegno, la prima modificazione che si realizza a carico del sistema cardiovascolare e' rappresentato dalla vasocostrizione dei distretti splancnici e dei centri motori encefalici. Tale fenomeno risponde ad un preciso finalismo: quello di spremere i serbatoi viscerali
di sangue onde aumentare la massa ematica circolante con conseguente maggior apporto di ossigeno in periferia.
I gruppi muscolari in attività, nello svolgere il loro lavoro, aumentano i processi metabolici locali con accumulo di acido lattico ed altri cataboliti acidi. Tali sostanze esercitano immediatamente un' azione di rilasciamento degli sfinteri precapillari chiusi, con successiva apertura alla profusione ematica dei relativi distretti capillari.
Contemporaneamente alla spremitura dei serbatoi ematici viscerali si ha un aumento del ritorno venoso al cuore, che si traduce in un aumento del volume telediastolico che comporta un allungamento delle fibre miocardiche nella fase presistolica, cioe' aumenta il PRECARICO (legge MAESTRINI-STARLING).
Aumentando quindi la forza contrattile e la velocita' di contrazione, si ha come conseguenza un aumento della gittata sistolica. Tale fenomeno deve tradursi in un aumento della pressione sistolica, sicchè aumenta la pressione differenziale, che costituisce uno dei più importanti meccanismi propulsori del sangue verso la periferia per intervento del recupero elastico delle grosse arterie, alla fine di ogni gittata sistolica.
L' aumento della pressione sistolica determina anche un maggior gradiente pressorio fra inizio del sistema vascolare ed il suo sfioccamento capillare.
Per effetto di ciò aumenta la velocità di flusso nelle arterie e nelle arteriole comportando un aumento delle resistenze di attrito. Contemporaneamente però cresce la superfice complessiva della sezione capillare perfusa, pertanto risulta complessivamente aumentato il volume di deflusso in periferia con automatica diminuzione delle resistenze periferiche.
La stessa velocità di flusso che aumenta nei vasi distributori, subisce un rallentamento invece nei distretti capillari, vasi nutrizionali, proprio per l' enorme dispiegamento del loro letto.
Ed ovviamente ciò risponde a precise esigenze fisiologiche: nei territori impegnati nel lavoro serve che il sangue arrivi presto ma serve anche che possa soffermarsi per il tempo necessario per gli scambi utili al proseguimento del lavoro stesso.
Il sangue dopo aver attraversato i distretti capillari viene nuovamente avviato alla pompa (cuore) attraverso il sistema venoso che subisce modificazioni alquanto semplici legate alla velocità di flusso dovuta alle giaà accennate modificazioni pressorie.
Alle modificazioni del sistema venoso refluo e' legata un' altra modificazione dell' apparato cardiovascolare: la variazione di FREQUENZA CARDIACA. Infatti con la vasocostrizione del distretto splancnico e la spremitura dei serbatoi viscerali, si determina un aumento del flusso sanguineo attraverso le vene cave e nell' atrio destro dove vengono stimolati dei recettori nervosi (barocettori) ivi allocati, che inducono un proporzionale aumento di frequenza cardiaca per smaltire piu' rapidamente il nuovo accresciuto afflusso ematico ( riflesso di BAINBRIDGE).
Appare evidente come la funzionalità e l' efficenza dell' apparato cardiovascolare possano rientrare, anzi esserne i fattori limitanti nella possibilità per un organismo di produrre lavoro muscolare, infatti fino a qual punto una persona può spingere il proprio lavoro muscolare ?
E' chiaro che il concetto rientra nei limiti del metabolismo, ma questo a sua volta, condizionato dal sistema di trasporto dell' ossigeno, comprende l' apparato cardiovascolare, l' apparato Respiratorio, e la capacità di trasporto dell' ossigeno da parte del sangue, poichè modeste sono le possibilità di un lavoro in condizione di anaerobiosi.
Allora, se le capacità metaboliche di un organismo sono dipendenti dalla quantità di ossigeno utilizzata per quelle esigenze, la misura del CONSUMO MASSIMO DI OSSIGENO (VO2max) ci darà il limite metabolico della capacità di lavoro muscolare di quell' organismo. Ma nella valutazione del VO2max è implicita la capacità funzionale dei vari fattori che compongono il sistema trasportatore di ossigeno.
Così in laboratorio si può vedere facilmente come il VO2max può aumentare fino a 20 volte il VO2 a riposo, mentre la portata cardiaca e la differenza artero-venosa di ossigeno possono aumentare fino a 3-5 volte i valori di base.
Ne deriva che i fattori limitanti la capacità di lavoro di un individuo sano vengano rappresentati proprio dalle prestazioni del cuore come pompa (portata e frequenza cardiaca) e dalla utilizzazione dell' ossigeno in periferia (differenza artero-venosa di ossigeno).
Ecco pertanto profilarsi il significato dell' allenamento di un soggetto normale e gli obiettivi che esso deve prefiggersi e che noi possiamo riassumere in questo modo:
OBIETTIVI METABOLICI
- migliore utilizzazione dell' ossigeno a livello mitocondriale
- maggiore attività degli enzimi ossidativi
- aumento della sintesi di glicogeno e trigliceridi nei muscoli più interessati all' attività lavorativa
OBIETTIVI EMODINAMICI
- incremento della portata cardiaca
- riduzione della frequenza cardiaca per i vari livelli di spesa energetica sottomassimale
- migliore circolazione capillare con piu' ampio dispiegamento dei letti capillari nei muscoli che lavorano
- aumento del VO2max ( per riduzione della frequenza cardiaca e incremento della gittata sistolica e della differenza artero-venosa in ossigeno ( quest' ultima grazie appunto alla maggiore capillarizzazione muscolare).
PARAMETRI CARDIOVASCOLARI E PRESTAZIONE SPORTIVA
Fino a questo punto abbiamo trattato l' argomento relativo all' apparato cardiovascolare sotto l'aspetto prettamente anatomo-fisiologico, ma quello che più interessa in una trattazione finalizzata alla prestazione sportiva sono tutte le argomentazioni di carattere pratico che devono necessariamente far parte del bagaglio culturale di un Preparatore Atletico che voglia definirsi tale. Parlare quindi di gittata sistolica, portata, ecc... in fondo per noi operatori di campo è solo un mero valore aggiunto; noi abbiamo a che fare con la prestazione e con i parametri empirici ad essa legati, dunque per noi l' apparato cardiovascolare deve potersi riassumere in numeri tangibili, tali da poter farci rendere conto effettivamente di quello che si sta facendo ed eventualmente correggere o supportare una scelta metodologica o una strategia di allenamento.
Parliamo dunque dei parametri fisiologici legati all' apparato cardiovascolare.
1) PRESSIONE ARTERIOSA
Si misura con lo sfigmomanometro in mm Hg, usando anche il fonendoscopio.
Valori fisiologici sono da 70-90 per la minima, e per la massima fino ai 120-140 indipendentemente dall' età.
Di importanza fondamentale nello sport e' la pressione differenziale che determina il gradiente pressorio aumentato durante l' attività fisica (come abbiamo spiegato prima).
La pressione arteriosa massima (sistolica) corrisponde alla pressione che la parete dell' arteria deve vincere per far defluire il sangue all' interno del lume, la pressione minima (diastolica) è la pressione costante cui le pareti delle arterie sono sottoposte.
2) FREQUENZA CARDIACA ( FC , HR )
Questo è il pararmetro più tangibile che riflette l' efficienza dell' Apparato Cardiovascolare. Di essa dobbiamo sapere che può essere modificata da agenti endogeni (scariche ormonali, adrenalina, sistema simpatico) e da fattori esogeni (attività fisica, ecc...).
Sappiamo per esmpio che all' aumentare dell' intensità del lavoro fisico essa incrementa, e tende ad incrementare fino ai limiti massimi che sono stabiliti fisiologicamente in correlazione negativa con l' età (fig. 7).

relazione esistente tra l'intensità del lavoro (indicato con il VO2 (consumo di ossigeno)) e la Frequenza Cardiaca
fig. 7
A tale proposito vorrei ricordare che la FCmax in un soggetto sano può essere teoricamente calcolata come segue :
a) FCmax = ( 220 - eta') +- 10
b) FCmax = 198 - 0,925*(eta' - 20) +- 10
Essendo questo un dato teorico deve essere preso con beneficio di inventario, comunque posso dirvi che l' esperienza acquisita mi porta a ritenere valida la teoria almeno nel 90% dei casi esaminati (sono molti !).Quando è possibile (nel caso di soggetti allenati) conviene verificare la FCmax con un test massimale da sforzo ( es. 400mt di corsa veloce).
3) CALCOLO DEL MASSIMO CONSUMO DI OSSIGENO ( VO2max )
Rappresenta l' espressione della massima potenza aerobica, ed e' considerato uno dei più importanti fattori funzionali dell' organismo. E' l' indice della massima potenzialità di lavoro aerobico di un organismo (quindi il massimo lavoro possibile con metabolismo ossidativo) e si raggiunge quando siamo alla massima frequenza cardiaca. In altre parole la massima energia aerobica ottenibile attraverso le ossidazioni dei substrati energetici (glucidi e lipidi).
La sua unità di misura e' ml O2 / minuto, o ancora meglio ml O2 / Kg / minuto.
Dal punto di vista legato a fattori esclusivamente circolatori il VO2 può essere determinato dall' equazione di Fick, dove
VO2 = HR * gitt. sist. * (conc. arteriosa O2 - conc. venosa O2)
Teoriocamente è possibile sapere qual' è il VO2max di un soggetto sano, applicando la formula di Wassermann :
MASCHI di peso corrispondente a <= 0,79 * altezza (cm) - 60,7
allora
VO2max = peso (Kg) * (50,72 - (0,372 * eta'))
altrimenti
VO2max = ((0,79 * altezza) - 60,7) * (50,72 - (0,372 * eta'))
FEMMINE di peso corrispondente a <= 0,65 * altezza (cm) - 42,8
allora
VO2max = ( 42,8 + peso(Kg)) * (22,78 - (0,17 * eta'))
altrimenti
VO2max = altezza(cm) * (14,81 - (0,11 * eta'))
Esso può essere misurato in maniera diretta, con strumenti sofisticati e con protocolli ben definiti (Bruke, ecc...) o anche in maniera indiretta, basato sulla linearità della funzione che intercorre tra frequenza cardiaca e consumo di ossigeno (Test di Asmussen, Astrand, Ryming):
FC = A + B*VO2
Il test di Astrand si effettua su nastro trasportatore.
Rodolfo Margaria (il fisiologo padre della fisiologia muscolare in Italia), invece ha elaborato una equazione con la quale si può determinare il VO2max ed è la seguente:
FCmax * (VO2" - Vo2') +(FC" x VO2') - (FC' x VO2")
VO2max = ---------------------------------------------------------------------
FC" - FC'
Cioè conoscendo il consumo di ossigeno per un determinato esercizio e rilevando la frequenza cardiaca corrispondente, si può tracciare una retta di equazione sopradescritta ed estrapolando alla FCmax si può conoscere il VO2max. Il test proposto da Margaria per determinare il VO2max è lo STEP su scalino alto dai 30 ai 50 cm a ritmo predeterminato (es. 60 e 100 cicli al minuto).
Siccome il costo in VO2 per Kg di peso è costante in tutti i soggetti normali che non abbiano limitazioni funzionali negli arti inferiori, conoscendo l' altezza dello scalino, il ritmo con cui si esegue il test, è possibile sapere il VO2 per Kg al minuto, trovando la FC in ogni esercizio facilmente si puo' calcolare il VO2max.
Questa equazione può essere anche utilizzata per calcolare il VO2max, partendo dal test di Conconi, sapendo che il consumo di ossigeno in un corridore che corra con una buona tecnica a velocità costante è dato dalla seguente formula:
VO2 per la corsa = (2,917 * v + 0,000617 v^3) mmlO2/Kg/minuto
dove v = velocità in Km/h.
Per trovare la velocità in Km/h partendo da un riscontro cronometrico su una distanza espressa in metri è possibile applicare la formula seguente :
Km/h = (3,6 * distanza in metri) / tempo in secondi
esempio 100mt percorsi in 12", allora:
(3,6 * 100)/12 = 29,999 Km/h
Esiste una stretta relazione quindi tra VO2max e la velocità massima di corsa in una competizione aerobica, infatti se consideriamo che il consumo di O2 non implicato nella corsa è di circa 6mmlO2/Kg/minuto ( un pò superiore al consumo di O2 a riposo che e' di 4mmlO2/Kg/minuto), e l' energia che proviene dalla scissione del glicogeno con produzione di Acido Lattico, che può essere paragonata a circa il VO2max in un minuto, possiamo applicare la formula seguente per il calcolo della massima distanza percorribile in un tempo t definito:
metri = 5 * (VO2max - 6) * t + 5*VO2max
Appare evidente che se togliamo l' apporto dell' energia anaerobica, possiamo calcolare la quantità di percorso possibile a spese dei soli processi ossidativi ( che in questo caso può essere vista come una sorta di soglia anaerobica):
metri = 5 * (VO2max - 6) * t
Diventa evidente che più breve è la prova aerobica (comunque al di sopra dei 3 minuti), più l' apporto della glicolisi diviene importante, ma qui siamo in un altro settore che tratteremo in un altro paragrafo.
FREQUENZA CARDIACA E ALLENAMENTO
La dinamica attraverso la quale la FC varia la sua ritmicità durante l' esercizio fisico, è già stata descritta precedentemente (maggiore massa circolante, barocettori, ecc.... ).
Si sa anche che un soggetto che si allena vede diminuire nel tempo i livelli di FC per i possibili lavori sottomassimali, cioè allo stesso livello di sforzo sottomassimale, nel tempo, la risposta cardiaca è inferiore rispetto all' inizio, con conseguente meno fatica per il cuore.
Il perchè si manifesti questo fenomeno trova spiegazione nella SINDROME Generale di Adattamento (SEYLE) che è alla base dell' ALLENAMENTO. Nel caso del cuore, sappiamo che al crescere della spesa energetica per effetto di un lavoro fisico, la FC tende a salire per compensare la aumentata richiesta di ossigeno.
Il cuore in questa situazione deve far aumentare la FC insieme alla Gittata Sistolica, in altri termini deve far aumentare la PORTATA CARDIACA.
Abbiamo detto che un ciclo cardiaco (sistole-diastole) dura mediamente a riposo circa 0,8 secondi di cui 0,6 sec. per la diastole e 0,2 sec.per la sistole.
In situazioni di riposo quindi il cuore ha il tempo necessario per nutrirsi (durante la diastole), riempirsi e svuotarsi. Ma quando la FC aumenta diminuisce sostanzialmente il tempo di diastole, quindi diminuisce il tempo in cui il cuore si irrora attraverso il circolo coronarico, e diminuisce anche la gittata sistolica in seguito al poco tempo a disposizione sia di riempimento sia di eiezione.
Conoscere quindi le risposte cardiache ai vari livelli di sforzo risulta oltremodo importante per l' allenatore al fine di indicare con una certa precisione quali debbano essere le esercitazioni idonee a far instaurare i fenomeni di adattamento.
Riguardo alla FC distinguiamo due livelli principali di adattamento:
1 - la FREQUENZA DI SOGLIA AEROBICA
2 - la FREQUENZA DI SOGLIA ANAEROBICA
Si raggiunge la FC di Soglia AEROBICA quando si instaura la condizione di massimo riempimento ventricolare con massimo volume di eiezione (gittata sistolica), situazione che si verifica circa al 60%-70% della FCmax.
A questo livello di FC si produce l' adattamento cardiaco minimo al lavoro muscolare, e al di sotto del quale non si produce alcun beneficio. Il metabolismo a questo livello di FC e' essenzialmente LIPIDICO (importante nel caso al lavoro aerobico voglia associarsi un presidio dimagrante).
Le formule per calcolare individualmente il livello di SOGLIA AEROBICA sono :
COOPER > FCmax * 0,6
KARVONEN > ((FCmax-FCriposo) * 0,6) + Fcriposo
Mentre questo metodo empirico per indicare la Soglia Aerobica può essere preso in considerazione senza grosse probabilità di errore, più complesso e' il metodo per individuare realmente la vera SOGLIA ANAEROBICA.
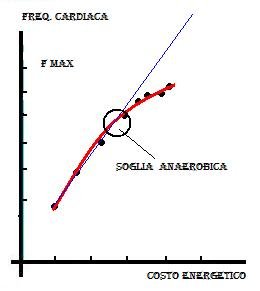
La Soglia Anaerobica rappresenta il limite massimo del metabolismo aerobico senza apprezzabile formazione di acido lattico (che comunque si forma anche a basssi livelli di lavoro muscolare, ma che viene metabolizzato dall' organismo senza accumulo),ed è rappresentata dalla FC che si raggiunge nel momento in cui nel sangue si trovano 4millimoli/litro di ACIDO LATTICO . In mancanza di strumentazione di analisi ematica, bisogna affidarsi a metodi indiretti come per esempio il test di CONCONI. Questo test si basa sulla osservazione della risposta cardiaca a livelli di forzo crescente (principio della linearità di relazione tra FC e Spesa energetica) (fig. 8).
report grafico del test di Conconi per la determinazione della soglia anaerobica
fig. 8
Mettendo su di un piano Cartesiano le variabili FC e Velocità di corsa (Spesa energetica), si nota che dopo le fasi iniziali in cui è rispettata la dipendenza lineare tra le due variabili, ad un certo livello di sforzo la dipendenza non è più lineare, quindi all' incremento di velocità non corrisponde più l' incremento corrispondente di FC. Si ha cioè una inclinazione della retta che quindi perde la linearità.
Il punto in cui si verifica questo fenomeno è individuato da due coordinate: una certa FC ed una certa Velocità. Questo punto, detto di DEFLESSIONE o IMPENNATA ANAEROBICA, è il punto della SOGLIA ANAEROBICA.
In mancanza della possibilità di effettuare il test, si può far riferimento, ma con tutti i benefici di inventario possibili, alle formule seguenti :
COOPER > FCmax * 0,85
KARVONEN > ((FCmax-FCriposo) * 0,85) + FC riposo
E’ possibile determinare la Soglia Anaerobica anche effettunado un altro semplice test da campo validato da Bisciotti e Sagnol dell'Università di Lione; il test consiste nel correre alla massima velocità possibile i 3000 metri e (uno o due giorni dopo) i 2000 metri registrando i tempi in secondi.
Viene fatta la differenza dei tempi ottenuti e la differenza tra le due distanze percorse, quindi si ottiene il tempo in cui vengono percorsi i 1000m (che sono la differenza tra 3000 e 2000 metri). Se per esempio si impiegano 11’30” per fare i 3000m e 8’ per i 2000m avremo : 730“ (che corrispondono a 11’30”) e 480” (che corrispondono a 8’) la cui differenza dà 250”. Quindi 1000m in 250” corrispondono a 4 m/s che rappresenta la "velocità critica" o velocità di soglia anaerobica (per avere la velocità espressa in Km/h basta moltiplicare 4m/s x 3,6 otterremo così 14,4 Km/h).
RESISTENZA LATTACIDA
Il protrarsi di uno sforzo di intensità massimale determinerà ineluttabilmente la produzione di Acido Lattico in quantità sempre più massicce fino all' esaurimento della capacità contrattile dei muscoli impegnati nello sforzo.
L' acido lattico, essendo un elemento tossico del muscolo che interviene sull' ambiente in cui il muscolo è chiamato ad operare, variandone sensibilmente il ph (valori < 7), determina l' inibizione biochimica sul sistema di accoppiamento acto-miosinico, con conseguente arresto progressivo della capacità di contrarsi dei singoli sarcomeri ed in ultima analisi del muscolo in toto.
Va da sè che l' obiettivo principale dell' adattamento che si deve perseguire nell’ allenamento, deve mirare al ritardo con cui il fenomeno suddescritto viene ad istaurarsi.
Le strade da percorrere, quindi, saranno indirizzate prevalentemente verso l' incremento, da parte del sistema biochimico degradatore, degli elementi necessari alla produzione di energia di resintesi, e , da parte del sistema muscolare, verso l'elevazione della soglia di sopportazione all' acido lattico.
La risposta fisiologica di quest'adattamento consisterà in un marcato aumento degli enzimi della catena glicolitica, un aumento dei depositi di glicogeno, ed un aumento delle sostanze bicarbonate che dovranno tamponare la comparsa dell' acido lattico.
Le metodiche allenanti che perseguono questi obiettivi sono da ricercarsi in quelle che prevedono prove ripetute ad intensità elevata intervallate da pause più o meno lunghe.
Se il fine è quello di far acquisire una maggiore riserva di glicogeno, quindi aumentare la Capacità del sistema, ci si porrà in condizione di eseguire movimenti di intensità sufficientemente elevata ma non massimale, per un periodo di tempo superiore al minuto, ripetendo il ciclo per 6 - 8 ripetizioni con pause incomplete della durata di minimo 5' fino ad un massimo di 8'- 10' (es. 6 rip. di 500m 80%max con 10' di rec.).
Se invece lo scopo dell' allenamento è quello di rendere più efficiente il sistema di erogazione energetico (POTENZA), i ritmi di lavoro saranno massimali o supermassimali e necessariamente i tempi di impegno muscolare saranno più brevi del periodo di competizione ma le velocità dovranno essere superiori; in questo caso le ripetute saranno di numero contenuto ed i tempi di pausa più lunghi, tali da consentire comunque un ripristino quasi totale (es. 4 volte i 300m 100% del massimale, recupero minimo 15').
RESISTENZA AEROBICA
Quando uno sforzo fisico si protrae da qualche minuto a qualche ora, siamo di fronte ad un fenomeno motorio che incentra la sua realizzazione sulla capacità soggettiva di produrre ed utilizzare energia proveniente prevalentemente dalle ossidazioni, cioè dalla degradazione metabolica dei substrati energetici per mezzo dell' ossigeno. Da qui l'attributo: Aerobica.
I sistemi ed apparati che questo fenomeno coinvolge sono molteplici, ed è per questo motivo che la resistenza aerobica è la qualità fisica più facilmente migliorabile in quanto risultante di adattamenti allargati a piuù elementi.
Cuore, polmoni, circolazione, respirazione, situazione intima muscolare, elementi sanguinei, organi cellulari, enzimi, depositi energetici, forza di volontà: tutto ciò ed altro è il riassunto della Resistenza aerobica.
Tutto ciò comunque e' facilmente allenabile.
Certo che per puntare a risultati di vertice, bisogna essere naturalmente dotati di prerequisiti già biologicamente determinati.
Come precedentemente specificato, anche in questo settore del movimento si distinguono le caratteristiche di Capacità e Potenza del sistema erogatore.
La Capacità del sistema aerobico e' teoricamente infinita in quanto il combustibile utilizzato è l'insieme dei depositi lipidici e glucidici apportati con l' alimentazione, quindi facilmente reintegrabili, e il comburente delle reazioni biochimiche è l' ossigeno tratto dall' aria che respiriamo, praticamente sempre disponibile.
L' adattamento di questa caratteristica va ricercata nella capacità dell' organismo di rendere disponibile una grande quantità di energia potenziale per mezzo degli accumuli di sostanze energetiche, e di rendere il più possibile agevoli gli scambi gassosi tra sangue arterioso e tessuti muscolari.
Il mezzo tecnico adatto allo scopo e' il lavoro continuativo e costante protratto per tempi più o meno lunghi, comunque dell' ordine dell' ora e a frequenze cardiache medio-basse.
Cosa avviene nell' organismo se seguiamo un piano di allenamento concepito come abbiamo detto ?
1) Il primo adattamento lo ritroviamo nella funzionalità cardiaca, intendendo per questa una migliore contrattilità del miocardio ed un marcato incremento della portata (gittata sistolica x frequenza cardiaca). A parità di sforzo, dopo un certo periodo di allenamento, la frequenza diminuisce con conseguente diminuzione dell' affaticamento del cuore.
2) Un aumento del numero dei capillari che si aprono a livello muscolare e centrale, con conseguente miglioramento dell' apporto di ossigeno ai distretti biomeccanici e al miocardio.
3) aumento del numero dei globuli rossi e dell' emoglobina.
4) Aumento dei mitocondri, con conseguente maggiorata produzione di energia aerobica di resintesi.
5) Diminuzione della parte adiposa della composizione corporea.
6) Miglior funzionamento della meccanica respiratoria.
Insomma un miglioramento della funzionalità organica che si traduce in un miglioramento della efficienza fisica generale e dello stato di salute. Non a caso anche il recupero funzionale del postinfartuato è costituito soprattutto da esercitazioni aerobiche di bassa intensità.
Solitamente si esegue un allenamento che consenta una frequenza tra il 60% e il 70% della frequenza massima per un tempo che varia da 1h a 2h; questo lavoro si chiama FONDO LENTO.
Cosa avviene invece se diminuiamo i tempi di applicazione ed aumentiamo la frequenza cardiaca durante l' allenamento ? In questo caso creeremmo i presupposti per migliorare la velocità delle reazioni ossidative e la capacità di resistere a ritmi elevati; praticamente incrementeremmo la POTENZA del sistema aerobico e i presupposti psicologici per affrontare una competizione.
Le metodiche allenanti sono molteplici e si differenziano a seconda dei ritmi di lavoro. Distinguiamo:
- FARTLEK (gioco di ritmi : piano-medio-piano-veloce ecc...)
- Fondo Medio (30'-45' frequenza cardiaca 85% fcmax)
- Fondo Veloce (15'-25' frequenza cardiaca 95%-105% fc di soglia)
- Interval Training (distanza, tempo, numero prove, pausa, attività di pausa)
- Ripetute a ritmo di gara.
Bibliografia
R. Margaria Fisiologia muscolare e meccanica del movimento. Biblioteca dell'Est 1973
A.A. vari : Fisiologia dell’uomo – Edi Ermes -2000
W. McArdle–E. Katch–V. Katch : Fisiologia applicata allo sport– C.E.Ambrosiana -2000
BrooksGA. Anaerobic treshold: rewiew of the concept and direction for future research. Med. Sci. Sports. Exerc. 17: 22-31, 1985.
P.G.J.M.Janssen Training, lactate, pulse rate. Polar Electro Oy 1989
Billat V. A method for determining lactate steady state whith two stages at submaximal worksload. Eur. J. Appl. Physiol. 69: 194-202, 1994.
Di Prampero PE., Fusi S., Antonutto G. Il concetto di soglia anaerobica. Medicina dello Sport. 51: 393-400, 1998.
Tanaka K., Matsuura Y., Kumagai S., Matsuzaka A., Hirakoba K., Asano K. Relationship of anerobic threshold and onset of blood lactate accumulation with endurance performance. Eur. J. Appl. Physiol. 52: 51-56, 1983.